
ABSTRACT Insulin resistance is an important
risk factor in the development of cardiovascular diseases such as hypertension
and atherosclerosis. However, despite its importance, the specific role of
insulin resistance in the etiology of these diseases is poorly understood. At
the same time, ethanol (ETOH) is a potent vasoconstrictor that primarily
induces down regulation of mitogen activated protein kinases (MAPKs) which
could exacerbate insulin resistance and possibly lead to cardiovascular
diseases. This article describes how chronic ETOH exposure interferes with
insulin signaling in hypertensive vascular smooth muscle cells (HVSMCs) which
leads to the alteration of MAPKs, the major signaling molecules. Elevated (50 -
800 mM) chronic exposure (24 hr) of HVSMCS to ETOH prior to insulin stimulation
decreased insulin-induced ERK 1/2 (MAPKs) and AKT expression. Similar
experiments were conducted using normotensive cells from rat. These cells
showed reductions in insulin-induced ERK 1/2 phosphorylation as well, but only
at higher concentrations of ETOH (400 - 800 mM). These alterations in insulin
signaling could provide an alternative molecular mechanism that may increase
the risk of insulin resistance, thus increasing the possibility of
cardiovascular diseases. Keywords: HVSMCs; Ethanol; Insulin; ERK 1/2; AKT 1.
Introduction Insulin resistance can lead to the development of cardiovascular
diseases such as hypertension and atherosclerosis [1-3]. However, there is
little information known about the role of insulin resistance in cardiovascular
diseases. As a result of chronic alcohol consumption, insulin resistance can
occur. Insulin release and activation is important in the body to alleviate the
outcome of diabetes and in some cases, hypertension [4,5]. Chronic ethanol
(ETOH) consumption can also affect insulin’s ability to bind to its receptor,
therefore, leading to insulin resistance [4,5]. However, there are not many
studies to prove this effect, especially in a hypertensive phenotype. Studies
have shown that moderate consumption of alcohol is not a precursor to coronary
artery disease, but chronic consumption of alcohol can be detrimental [5].
Insulin signaling is important in the cell for the release of insulin to
control glucose intake and lipid metabolism [5]. Insulin has an important role
in the biological process because it binds to its receptor (insulin receptor)
to initiate glucose metabolism [5]. The insulin receptor is a transmembrane
domain spanning tyrosine kinase receptor composed of alpha and beta subunits
that mediate the actions of insulin [6-8]. Insulin is closely associated with
hypertension, non-insulin dependent diabetes, atherosclerosis, and dyslipidemia
[6,7]. As a result, insulin activation of PI3K-AKT (Phosphoinositide 3-Kinase),
which mediates neuronal survival, motility, energy metabolism, and plasticity
is impaired [6,8,9]. The extracellular signal-regulated kinases (ERKs), also
known as extracellular mitogen activated protein kinases (MAPKs), have been
found to be altered by ethanol treatment of vascular smooth muscle cells from the
aorta of a rat [5,9-11]. This effect of ETOH has been shown to be manifested
via several pathways by the use of signaling inhibitors, such as PKC, leading
to a cascading effect in treated cells [11,12]. Several studies have shown that
insulin activates a complex set of intracellular responses, including the
activation of mitogen- S. D. WILLIAMS, B. WASHINGTON 187 activated protein
kinases ERK 1/2 [13] and AKT [5]. The normal role of AKT in the cell is to
propagate insulin receptor signaling to downstream effectors [14]. AKT is
downstream of PI3K, both of which are a part of one of the major pathways in
insulin signaling. The other major pathway is the mitogen activated protein
kinase pathway. It is also known that AKT has a role in activating insulin response
[10]. Alteration of AKT expression is exhibited by ETOH impaired insulin
signaling in the body and a decrease in glucose transport of rat cardiac muscle
cells [10]. Although insulin response can be altered with chronic ETOH, there
are other factors that can alter this response such as genetics and the
person’s environment [5]. Chronic ETOH consumption in experimental animal
models has been shown to alter insulin signaling events via the mitogen
activated protein kinases producing insulin resistance in the liver. These
adverse effects of ethanol have been shown to be the result of the inhibition
of insulin or insulin-like growth factor which alters mRNA and DNA synthesis
and the activation of proapoptotic signals through PI3K and AKT [15-17].
Results from previous studies suggest that ETOH impairment of insulin action is
likely to be downstream from PI3K, however, the mechanisms underlying the
effects of ETOH on insulin resistance and the effect of insulin resistance on
the development of cardiovascular diseases remain to be determined [5,18]. This
paper describes how chronic ETOH exposure can alter insulin signaling in HVSMCs
using mitogen protein kinases as indicators. Chronic ETOH exposure’s effect on
insulin signaling has not been analyzed before in hypertensive cells. 2.
Materials and Methods 2.1. Reagents and Antibodies ERK 1/2 (p44/p42) and AKT
antibodies were purchased from Cell Signaling (Beverly, MA); anti-rabbit IgG
antibodies (horseradish peroxidase linked) from Amersham Bioscience
(Piscataway, NJ); and ECL detection system was obtained from Pierce
Biotechnology (Rockford, IL). Other supplies include Dulbecco’s Modified
Eagle’s Medium (Amersham), fetal bovine calf serum, penicillin and streptomycin
were purchased from Sigma/Aldrich (St. Louis, MO). 2.2. Cell Culture Vascular
smooth muscle cells (VSMCs) were received from Vanderbilt University. Cells
were cultured in DMEM containing 10% fetal bovine serum, 2% penicillin and
streptomycin. Subcultured passages were between 3 and 12. Cells were maintained
at a pH of 7.1 in 75 cm2 flasks under a humidified atmosphere of 5% CO2, 95% O2
at 37˚C and plated in 6-well falcon plates. 2.3. Insulin Treatment For dose
response experiments, cells were stimulated with 1, 2, 4, 8, and 16 μM insulin
for 30 min. Insulin was aspirated from each well. For time course experiments,
cells were stimulated with 8 μM of insulin for 1, 5 10, 20, and 40 min. Cells
were lysed with 300 - 500 μl Laemmli Sample Buffer (2% SDS, 25% glycerol, 0.01%
bromophenol blue, and 62.5 mM Tris-HCl pH 6.8). Cells were then scraped from
the monolayer surface and collected in microcentrifuge tubes. 2.4. Ethanol
Treatment Hypertensive and normal rat cells were induced with 50, 100, 200,
400, and 800 mM ETOH. Control cells were induced with a DMEM (-) solution
containing no serum that aids in cell proliferation. After a 24 hr incubation
period, ETOH was aspirated from all wells and cells (except controls) were
stimulated with 8 μM of insulin. Insulin was aspirated from each well and the
cells were lysed with 300 - 500 μl Laemmli Sample Buffer (2% SDS, 25% glycerol,
0.01% bromophenol blue, and 62.5 mM Tris-HCl pH 6.8). Cells were then scraped
from the monolayer surface and collected in microcentrifuge tubes. 2.5. Western
Blot Analysis Whole cell lysates were collected and diluted with sample buffer
to equal concentrations of 40 μg/20 μl. Lowry protein assay was conducted to
determine standard protein concentration. Protein samples were then separated
along with rainbow markers to measure the molecular weight of proteins on a 10%
SDS-polyacrylamide gel from Bio-Rad Laboratories (Hercules, CA) at 200 volts
for approximately 50 min. Proteins were then transferred to a nitrocellulose
membrane from Amersham Biosciences (Piscataway, NJ) using a semi-dry transfer
apparatus at 10 volts for 90 min. Blots were blocked with 2% non-fat dry milk
in TBS (Tris-Buffered Saline) for at least one hour. After which blots were
incubated with primary antibodies ERK 1/2 or AKT overnight followed by
anti-rabbit secondary antibody for 1 hour. Blots were immersed in
chemiluminscent solution and developed in a dark room. 2.6. Statistical
Analysis All experiments were performed in triplicate and expressed as means ±
SE of the density using arbitrary units from three individual experiments.
Statistical significance was determined with paired or unpaired onetailed
Student’s t-test, with P < 0.05 considered significant. Open Access CellBio
188 S. D. WILLIAMS, B. WASHINGTON 3. Results 3.1. Insulin Induction Increases
ERK 1/2 and AKT Expression in Hypertensive VSMCs Before we could determine the
effect of ETOH on insulin signaling, we first had to determine the maximum
concentration of insulin it would take to stimulate phosphorylation of ERK 1/2
and AKT via Western Blotting analysis. HVSMCs were stimulated with a
concentration range of 1 - 16 μM of insulin for 30 min. Figure 1 denotes that
stimulating HVSMCs with 1 - 16 μM of insulin, increased ERK 1/2 phosphorylation
by approximately 23%. In addition, this insulin stimulation increased in AKT
expression 33% above basal with a maximum expression detected with 8 μM of
insulin (Figure 2). This data suggests that AKT activation in HVSMCs is more
sensitive than ERK 1/2 to insulin signaling. 3.2. Insulin Induction Increases
ERK 1/2 and AKT Expression in Normal VSMCs In order to determine the effect of
insulin signaling in normal cells, VSMCs were stimulated with a concentration
range of 1 - 16 μM of insulin. Maximal expression of phosphorylated ERK 1/2
occurred with 8 μM of insu- (a) (b) Figure 1. Dose response curve for insulin
on ERK 1/2 and AKT expression in HVSMCs. Cells were stimulated with 1, 2, 4, 8,
and 16 μM insulin for 30 min and lysate harvested. (a) Western Blot Analysis of
SHR (Spontaneously Hypertensive) lysate probed with antibodies for ERK 1/2 and
AKT expression; (b) Graphical representation of data by densitometry analysis
software was taken from a mean of three experiments p < 0.05 compared to
control. (a) (b) Figure 2. Dose response curve for insulin on ERK 1/2 and AKT
expression in normal VSMCs. Cells were stimulated with 1, 2, 4, 8, and 16 μM
insulin for 30 min and lysate harvested. (a) Western Blot Analysis of WKY
(Wistar Kyoto) lysate probed with antibodies for ERK 1/2 and AKT expression;
(b) Graphical representation of data by densitometry analysis software was
taken from a mean of three experiments p < 0.05 compared to control. lin,
which was significantly different from basal (Figure 2). No significant
increases in ERK 1/2 were observed between 1 - 4 mM (Figure 2). Similar experiments
showed that AKT expression was significantly increased throughout all
concentrations except 1 μM with maximal expression at 8 μM. Insulin seemingly,
also induced increases in AKT with 1 - 8 μM in normal VSMCs (Figure 2). This
increase in expression of AKT was not observed when cells were stimulated with
16 μM insulin which resulted in complete inhibition. 3.3. Chronic ETOH Impairs
Insulin Signaling in Hypertensive VSMCs In order to investigate whether ETOH
alters insulin signaling in HVSMCs, cells were treated chronically (24 hrs)
with 50 - 800 mM of ETOH. After ETOH treatment HVSMCs were stimulated with 8 μM
insulin for 30 min and Western Blotting was performed. As a result, ERK 1/2
expression significantly decreased with 50 - 800 mM ETOH treatment by 10%
compared to insulin stimulation only (Figure 3). Using the same treatment range
(50 - 800 mM) AKT expression was evaluated by chronic ETOH treatment. After 24
hours ETOH treatment, AKT expression HVSMCs significantly decreased (Figure 3).
This decrease was gradual as the concentration increased. Open Access CellBio
S. D. WILLIAMS, B. WASHINGTON 189 (a) (b) Figure 3. Twenty-four hour ETOH
treatment of HVSMCs. Cells were treated with specified concentrations of ETOH
(50 - 800 mM) and stimulated with 8 μM insulin for 30 min and lysate collected.
(a) Western Blot Analysis of SHR (Spontaneously Hypertensive) lysate probed
with antibodies for ERK 1/2 and AKT expression; (b) Graphical representation of
data by densitometry analysis software was taken from a mean of three
experiments p < 0.05 compared to control. 3.4. Chronic ETOH Impairs Insulin
Signaling in Normal VSMCs In order to investigate whether ETOH alters insulin
signaling in normal VSMCs, we evaluated chronic ETOH treatment in them as well.
Cells were treated chronically with 50 - 800 mM of ETOH for 24 hr. After ETOH
treatment, VSMCs were stimulated with 8 μM insulin for 30 min. Fifty and 100 mM
of ETOH had no significant effect on insulin-induced AKT expression when
compared to insulin stimulation alone. However, 200 - 800 mM reduced
insulin-induced AKT expression approximately 20%. Chronic ETOH treatment of
normal VSMCs seems to cause an increase in signal at 50 mM and a decrease at
higher concentrations when compared to the insulin induced increase alone
(Figure 4). 4. Discussion The research findings in this paper provide evidence
for changes in mitogen-activated protein kinases possibly contributing to the
onset of cardiovascular disease. The focus of this study is to show the
association of chronic ethanol-induced changes in ERK 1/2 and AKT expression in
HVSMCs. Recent reports show that hypertensive (a) (b) Figure 4. Twenty-four
hour ETOH treatment of normal VSMCs. Cells were treated with specified
concentrations of ETOH (50 - 800 mM) and stimulated with 8 μM insulin for 30
min and lysate collected. (a) Western Blot Analysis of WKY (Wistar Kyoto)
lysate probed with antibodies for ERK 1/2 and AKT expression; (b) Graphical
representation of data by densitometry analysis software was taken from a mean
of three experiments p < 0.05 compared to control. persons are predisposed
to the development of diabetes. [19]. Seventy-five percent of cardiovascular
diseases are attributed to diabetes, hypertension, and alcohol, indicating that
this is a contributing factor to the onset of hypertension. In Figure 1, ERK
1/2 phosphorylation was increased in hypertensive cells with increasing
concentrations of insulin. This confirms that cells are able to survive with
higher concentrations of insulin indicated by ERK 1/2. On the other hand, when
AKT stimulation occurs, it has been reported that cellular expression
decreases. This could be due to the alteration of insulin signaling. In Figure
2, normal VSMCs with ERK 1/2 and AKT decreased with 1 - 8 μM insulin. With 16 μM
of insulin, both ERK 1/2 and AKT expression was inhibited. In Figure 3,
hypertensive cells treated with ethanol ERK 1/2 and AKT both decrease in
expression as the concentrations of ETOH increase which could mean that ethanol
is affecting insulin signaling. Figure 4 depicts a biphasic effect of ETOH on
insulin signaling in normal cells which suggest that there may be different
effect in normal cells. Cardiovascular diseases include atherosclerosis and
hypertension. Risk factors for such diseases of the cardiovascular system
include family history, diabetes, obeOpen Access CellBio 190 S. D. WILLIAMS, B.
WASHINGTON sity, smoking, excessive alcohol intake, and a diet high in salt
and/or low in antioxidant nutrients. Individuals with hypertension are at
increased risk for atherosclerotic diseases such as stroke, heart, and kidney
disease which can be exacerbated by diabetes and alcohol [20]. The adverse
effects of long-term excessive use of alcohol are similar to those seen with
other sedative-hypnotics drugs (apart from organ toxicity which is much more
problematic with alcohol). Though the underlying mechanisms remain undefined,
accumulating evidence strongly suggests that ETOH interferes with insulin’s
action by altering mitogen activated protein kinases. The major signaling
molecules, MAPKs, implicated in the biological actions of insulin, and the
expression of the insulin receptor may be major factors leading to
cardiovascular diseases. 5. Acknowledgements The authors would like to
acknowledge The Research and Engineering Training Program (REAP) and Title III
Program for providing support for this work.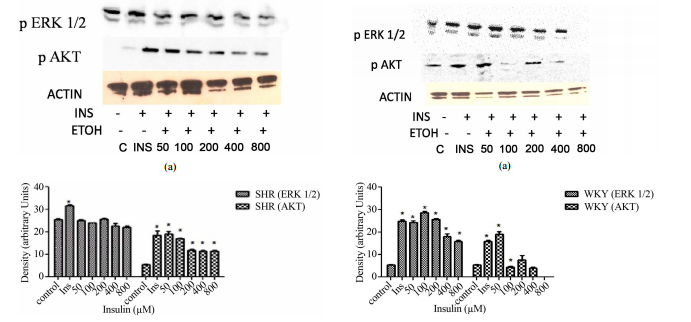








论文发表 是一个专门从事期刊推广、论文发表、论文发表辅导的机构。融合收集数百家期刊杂志社征稿评职称,发表论文,以供广大作者免费阅览,以期待各位作者在短时间内掌握每种期刊征稿要求、审稿范围,在日常繁重的工作中短时间开心、放心快速投稿、在适合的时间拿到刊物,评上职称,不为挤不上评职称的末班车而烦恼。
本站主要整合如下评职称,发表论文:
教育论文发表
经济论文发表
科技论文发表
医学论文发表
计算机论文发表
文学论文发表
农业论文发表
学报论文发表
其它论文发表等,另期待更多的杂志编辑与本站联系文:
85782530(普刊发表)
82534308(核心发表)共同以正期刊界正刊之气,维护学术良好的氛围。
免责声明:本网所提供的信息资源如有侵权、违规,请及时告知!
论文网版权所有, 本站提供论文发表 论文投稿 发表论文 发表文章
文章只代表作者观点,并不意味着本站认同,部分作品系转载,版权归原作者或相应的机构;若某篇作品侵犯您的权利,请来信告知:lunww@126.com
中国论文网全权所属国际域名:www.lunww.com 中文域名:论文发表 技术支持:论文发表 国家双软认定单位 法律顾问:全民安律师事务所
期刊合作加盟咨询:5715378(加盟、投诉、建议) 论文发表咨询电话:18262951856 论文发表投稿邮箱:lunww@126.com
Powered by 论文发表 © 2010-2020

苏ICP备19023845号-4













